| Acta Bioquímica Clínica Latinoamericana |
| Proteasas y cáncer
Estas enzimas desempeñan una función esencial en el cáncer, porque promueven el crecimiento de los tumores y fomentan la formación de metástasis. El Comité de Redacción de Acta Bioquímica Clínica Latinoamericana ha adaptado este artículo aparecido en ?Investigación y Ciencia?, mayo de 2000, por considerar de interés su difusión a través del FABA-Informa. |
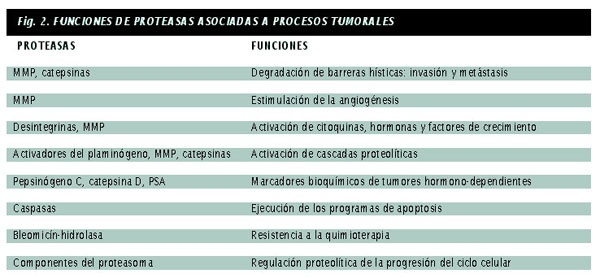
| Las proteasas son enzimas proteolíticas. Degradan total o parcialmente las proteínas. Y pueden hacerlo porque hidrolizan los enlaces que mantienen unidos entre sí a los aminoácidos, los bloques moleculares que construyen las proteínas de todos los organismos vivos. Al imaginar una vida sin proteasas, lo primero que viene a la mente es una sensación de indigestión: las proteínas de la dieta no se degradarían en el estómago y no se podría obtener energía para sobrevivir. Esta función digestiva de las proteasas contribuyó, hace más de 150 años, a la identificación y caracterización de estas proteínas en el jugo gástrico de animales de experimentación. Con el tiempo se ha ido conociendo mejor la significación biológica de las proteasas. Se les concede un papel central en la coagulación sanguínea y la cicatrización de heridas, la ovulación y la fertilización, la implantación y el desarrollo de los embriones, la formación de los huesos, la adquisición de recuerdos o la ejecución de los programas de apoptosis o suicidio celular. Tan impresionante diversidad funcional fue adquiriéndose a lo largo de miles de años de evolución biológica. Las proteasas involucradas en todos estos procesos actúan mediante sistemas en cascada, se combinan de manera muy diferente y multiplican en consecuencia sus efectos biológicos. Dado su enorme potencial destructor, se supone que la actividad enzimática de las proteasas se halla sujeta a una estricta regulación jerárquica. Cuando los mecanismos de control se alteran y las proteasas se forman en exceso, a destiempo o donde no procede, aparecen la artritis, la aterosclerosis, el cáncer y otros procesos patológicos similares. Proteasas y metástasis Por ello, parte de la investigación sobre el cáncer se centra en los mecanismos moleculares que conducen al desarrollo de metástasis. El proceso de formación de metástasis, un fenómeno muy complejo, consta de una serie de etapas secuenciales que implica la interacción entre células tumorales y componentes específicos de la matriz extracelular, la destrucción local de esa matriz y la migración activa de las células cancerosas a sitios distantes. Metaloproteasas de matriz extracelular Las MMP se reparten en cuatro subfamilias: colagenasas, gelatinasas, estromalisinas y metaloproteasas de membrana (MT-MMP). Las colagenasas rompen la triple hélice de los colágenos fibrilares; las gelatinasas degradan los colágenos de tipo IV y el colágeno desnaturalizado; pueden actuar de forma sinérgica con las colagenasas en la degradación de colágenos fibrilares. Las estromalisinas destruyen muchas proteínas extracelulares, incluidos los proteoglicanos, la laminina y la fibronectina. Finalmente, las MT-MMP tienen un espectro de acción muy restringido y hasta el momento sólo se conoce su participación en la activación proteolítica de otras metaloproteasas de matriz extracelular. La extraordinaria capacidad de las MMP para degradar los componentes de la matriz extracelular a pH fisiológico convertía a estas enzimas en candidatas ideales para intervenir en la destrucción hística que acompaña al cáncer. En los años sesenta se comenzó a estudiar esa posibilidad. El grupo encabezado por Lance Liotta, del Instituto Nacional del Cáncer en Bethesda, detectó la presencia de potentes actividades colagenolíticas en tumores epiteliales y mesenquimales. Observó que las células metastásicas de un carcinoma murino mostraban una mayor capacidad para degradar el colágeno que las células del tumor primario. Al propio tiempo se describió la síntesis de colagenasas por líneas celulares de osteosarcomas y carcinomas mamarios humanos, cuya actividad era bloqueada por una proteína de cartílago, un tejido resistente a la invasión por células neoplásicas. Pero no tardó en comprobarse cuán duro y complejo era el problema. El número de proteasas asociadas al cáncer superaba el esperado, justificable tal vez por la diversidad de sustratos a degradar por esas enzimas. Además, y pese a que las MMP parecían ser las proteasas idóneas para desempeñar estas funciones (por la naturaleza de sus sustratos y por el pH neutro al que actúan), comenzó a sugerirse la intervención de nuevas proteasas en el desarrollo del cáncer. Se abría el abanico. Al estudio de las metaloproteasas se sumó el de las cisteín-proteasas -catepsinas B y L-, el de las serín-proteasas -activadores del plasminógeno- y el de las aspartil-proteasas, como la catepsina D. Señales moleculares El humor maligno Desde una perspectiva actual, el desequilibrio humoral equivaldría a la pérdida del equilibrio entre los niveles de proteasas activadas y de sus inhibidores endógenos, causante de la destrucción de la matriz extracelular y, por ende, de la progresión tumoral. El ?humor maligno? de la medicina clásica estaría, visto hoy, constituido mayoritariamente por proteasas como la estromalisina-3 o la colagenasa-3, que se concentran en las zonas de invasividad tumoral. La colagenasa-3 Al estudiar la naturaleza de las señales moleculares enviadas por los tumores a las células normales con el fin de que éstas sinteticen la enzima, se han identificado varios factores solubles capaces de estimular la producción de la colagenasa-3. De éstos, la interleuquina-1 y el factor de crecimiento TGF-b, una vez sintetizados y secretados por las células epiteliales tumorales, inducen la expresión de la colagenasa-3 en las células estromales inmediatamente adyacentes al tumor. Tras la síntesis y activación de esta enzima a través de una cascada proteolítica, en la que intervienen las metaloproteasas MT1-MMP y la gelatinasa A, la colagenasa-3 participaría en la destrucción hística que acompaña a la progresión tumoral. Se han venido recogiendo pruebas que vinculan la colagenasa-3 con la diseminación tumoral. Se ha observado una estrecha relación entre la concentración intratumoral de colagenasa-3 y la evolución de la enfermedad. La expresión de colagenasa-3 está asociada con una menor supervivencia global de las pacientes con cáncer de mama, lo que constituye un indicio indirecto de la participación de la enzima en la progresión tumoral. Resultados similares se han obtenido en carcinomas escamosos de laringe, condrosarcomas, carcinomas basocelulares de piel o diversos tumores de origen ginecológico, patologías que sobreexpresan la colagenasa-3. En todos los casos, la producción de la proteasa va asociada a una mayor agresividad local de los tumores, aunque varíen los mecanismos inductores de la síntesis enzimática según el tipo de tumor. En carcinomas de mama, los factores capaces de inducir la expresión serían el TGF-b o la IL-1, mientras que en condrosarcomas el bFGF tendría un efecto preeminente y en carcinomas de laringe actuarían el TNFa- y el TGF-a. En síntesis, la colagenasa-3 cumple los requisitos que cabía esperar de una proteasa implicada en el cáncer. Se trata de una enzima dotada de una actividad proteolítica diversa y potente, sintetizada en lugares donde no se exige su presencia y cuya capacidad destructora llega a degradar las proteínas que constituyen las barreras hísticas que tratan de impedir la progresión tumoral. Proteasas, apoptosis y angiogénesis Puesto que la regulación del ciclo celular y la apoptosis tienen que ver con el cáncer, parece obvio que las alteraciones de las proteasas implicadas en ambos procesos pueden facilitar la oncogénesis. Las proteasas pueden, además, instar la angiogénesis o formación de nuevos vasos sanguíneos, imprescindible para el suministro de nutrientes a los tumores. Proteasas y progresión tumoral En cualquier caso, se puede afirmar que ninguna proteasa, por sí sola, activa las propiedades invasivas y metastásicas de todos los tumores; antes bien, cada tipo de tumor utiliza distintas combinaciones de proteasas para facilitar su progresión. Así, además de metaloproteasas como la colagenasa-3 o la estromalisina-3, enzimas de otras clases como los activadores del plasminógeno o las catepsinas contribuyen en mayor o menor medida en el proceso de destrucción hística consustancial al desarrollo del cáncer. Tampoco la actividad proteolítica directa sobre la matriz extracelular agota la vinculación de las proteasas con los tumores dado que, además, son enzimas que estimulan la migración celular, modulan la angiogénesis tumoral, actúan como factores mitogénicos y confieren resistencia a la quimioterapia. En general, los niveles intratumorales de proteasas guardan correlación con una menor supervivencia global de los pacientes y con un menor intervalo libre de enfermedad. Por ello, se han introducido ya en clínica activadores del plasminógeno y ciertas MMP, como factores bioquímicos para ayudar a predecir la evolución tumoral. La importancia de los inhibidores de proteasas, que impiden la progresión de los tumores malignos, ha sido apreciada por la propia naturaleza al dotar a los organismos de una amplia batería de inhibidores endógenos. Existen inhibidores específicos de las distintas subfamilias de proteasas: serpinas, que bloquean la actividad de las serín-proteasas; cistatinas, que actuan sobre las cisteín-proteasas, o los TIMP, inhibidores hísticos de metaloproteasas. Se ha observado que la adición de TIMP a cultivos de células de melanoma o fibrosarcoma bloquea la capacidad invasiva de estas células, tanto en sistemas in vitro como in vivo. Y se ha demostrado que la sobreexpresión de estos inhibidores en fibroblastos transformados con el oncogén ras, reduce la velocidad de crecimiento de los tumores e inhibe su capacidad invasiva. Finalmente, la supresión de la expresión del TIMP-1 en fibroblastos murinos, mediante el empleo de ARN anti-sentido, confiere a estas células propiedades invasivas, tumorigénicas y metastásicas. Todas estas observaciones han estimulado el diseño de inhibidores sintéticos que remeden la operación de inhibidores endógenos. Se ha sintetizado una primera generación de inhibidores de MMP, algunos de los cuales ya se están utilizando en ensayos clínicos de tratamiento de tumores. Terapias antitumorales. |
{kind=link}